Home Page || Public Bookmarks || The Calvin Bookshelf
The Cerebral Symphony
Seashore Reflections on the
Structure of Consciousness
Copyright ©1989 by William H. Calvin. You may download this for personal reading but may not redistribute or archive without permission (exception: teachers should feel free to print out a chapter and photocopy it for students).
| |
7
Who Speaks from the Cerebral Cortex?
The Problem of Subconscious Committees
The map is not the territory.
the early semiotician Alfred Korzybski, 1933
[Jorge Luis Borges] talked of a country that prided itself on its cartographical
institute and the excellence of its maps. As the years went by, this institute would
draw maps of greater and greater accuracy until at last the institute achieved the
ultimate, the full-scale map. And, Borges says, if you wander through the desert
today, you can see places where portions of the map are still pegged to the region
they represent!
The point of all this, of course, is that our job as cognitive scientists is only to
chart the territory of mental life to establish the major phenomena and their
relationships, not to provide the full-scale map, not to replace a life richly lived
by the running of some computer program.... Our job as scientists, irrespective of
our job as philosophers, is twofold: not only to provide explicit accounts where
we can, but also to understand the limitations of those accounts. And so we must
always exist in that tension, between the uncharted and the unknown.
the American brain theorist
Michael A. Arbib
, 1985
Is the road to consciousness paved with Mexican hats? Or at least lateral inhibition? Probably,
as lateral inhibition is all about competition between adjacent cells for dominance, all about
finding maxima and minima -- and so it provides a way of judging the best choice among
alternatives. But some other properties of cells in the aggregate need to be mentioned before
returning to consciousness per se -- such as sensory "maps" in the cerebral cortex.
Each cell in the visual pathways has a receptive field, one of whose properties is a "center" -- a
point in space toward which that cell's attention seems focused. Its neighboring cells are usually
focused on about the same spot -- but there is a drift, so that cells farther away seem to
concentrate upon more distant points in space. This drift is generally orderly, so we can make
maps, e.g., simplifications that plot only the centers. Cortical maps tend to be distorted because
the cells are more interested in some things than others -- rather like those maps of the world that
show the continents resized according to their population, or gross national product.
THE SUNRISE WAS HAZY THIS MORNING, thanks to all the humidity of the sea air. Since
the sun rises in the northeast in midsummer, it is over Cape Cod, as seen from Woods Hole,
rather than hanging over the Atlantic Ocean as you'd expect. The Cape is shaped somewhat like
the arm of a strongman flexing his biceps. The cocked wrist out at the northeast end is where
Wellfleet and Truro are, with Provincetown the clenched fist. Eastham is on the forearm,
Chatham is on the south side of the elbow looking down toward Nantucket. Hyannis is about the
triceps, Dennis the biceps. The armpit would correspond to Falmouth and Woods Hole, and the
head would be up along Cape Cod Bay about Plymouth Rock (where the Pilgrims landed),
towered over by Boston and Cambridge, as befits their institutions of higher learning.
The armpit image of Woods Hole might have once been accurate, back in the days before the
Marine Biological Laboratory and the Woods Hole Oceanographic Institute were founded. In the
late nineteenth century, Woods Hole was industrial and a railroad terminus for the train ferries
between Boston and New York (the great parking lot for the Vineyard and Nantucket ferries was
once a railroad switching yard, and a pall of coal smoke often hung over it).
But the most potent smell came from a fertilizer factory that mixed local fish called menhaden
with bat guano mined from cave floors in the tropics. Nine whaling ships operated out of this
port between 1815 and 1860, and they smell pretty bad too. The Pacific Guano Company went
out of business in 1880; the whaling ground to a halt because of greed (they foolishly helped to
exterminate entire species of whales).
The stench may be gone now, but Woods Hole isn't exactly the virginal salt marsh that it used to
be. Earth-moving machinery has remade its landscape, sculpting the fill dirt into a bulldozer
operator's idea of landscape aesthetics. Too bad they didn't straighten out those diabolical
twisting roads while they were at it, though half of the blind corners could be fixed with hedge
clippers. Perhaps blind corners are the local equivalent of a hair shirt.
The shape of Cape Cod suggests that it might have been the terminal moraine of an ice-age
glacier. Those of us who live in glacier country get used to seeing moraines. Hiking around
Mount Rainier each summer, one sees glaciers melting back, exposing the big piles of rubble that
they had plowed in front of them as they advanced. If you want to hike up to inspect the snout of
a retreating glacier, you will likely climb up over a series of tongue-shaped "terminal moraines"
on the way up the valley, recording the fitful advances and withdrawals of the snout.
Cape Cod is also tongue-shaped, and glaciers came down this far south (indeed, others made it to
Central Park in New York City, where one can see the parallel scrape marks made by rocks
trapped under the advancing front of the glaciers; Long Island is another moraine). The lowered
sea level during the glacial periods (as much as 30-40 stories) exposed a lot of the Continental
Shelf off the Atlantic coastline to habitation by land-loving glaciers.
The New England fishermen are always complaining about another leftover of the ice-age melt-off: the shallow bottom around the Continental Shelf is littered with big boulders, that ensnare
nets and lobster pots. They're the submerged equivalent of the "glacial erratics" which New
England farmers try to clear out of their fields, using them to build rocky fences. They were
carried along by the glacier, but the melting ice dropped them on the spot. And in the case of
those at lower elevations, the rising ocean eventually covered them.
There may have been a series of terminal moraines south of Boston. The Nantucket Shoals,
where the Atlantic Ocean is only one story deep in places, may represent the southernmost rubble
pile. The offshore islands of Martha's Vineyard and Nantucket, and the shallow waters in
between them, may be the terminal moraine of an advance that followed the meltback from the
shoals. Cape Cod itself may record, in part, the shape of the tongue of the glacier during its last
advance. One might expect that the next ice age will rearrange all this scenery, another glacier
plowing it all into a new terminal moraine. It's literally a case of sic transit gloria mundi.
THE BIKE TRAIL follows along the shores of Vineyard Sound from near Nobska all of the way
past Oyster Pond and into Falmouth. The Shining Sea Bikeway is an old railroad right-of-way
reclaimed for the public; now, you can now walk or bicycle nearly six kilometers without having
a car sideswipe you, once you get away from the ferry parking lot at Woods Hole.
Most people walk, though some masochistic runners are also in evidence. Bicycles come in all
flavors, but mostly this is a slow lane, not a fast one; people look around at the flowers, the
forest. Little paths lead off through the brush here and there. Then a pond pops into view,
complete with a pair of majestic swans.
One long stretch of the trail borders the ocean, and you can look south between Martha's
Vineyard and Nantucket to the open sea. And this sea really does shine; since you look to the
south and the sun is in the southern sky above it, a lot of sunlight gets reflected back into your
eyes. Katharine Lee Bates, whose home in Falmouth is a National Historic Something
hereabouts, wrote "America the Beautiful." Every time that I hear that lovely phrase "From sea
to shining sea" commemorating the East and West Coasts (where the sea only shines at sunrise or
sunset), I think of this south-facing beach where the sea shines all day.
Your view is unfortunately interrupted by the most obnoxious utility line yet seen. This is a
trunk line of some sort, tennis-ball thickness in most places -- more a sagging pipe than a wire.
Every few poles, it expands to football fatness, the tumors representing splices where a break has
been repaired. A pipe in the sky. It is so heavy as to require utility poles almost as closely
spaced as fence posts in places. And these utility poles march right along the beach, their guy
wires causing you to detour as you walk along near the lapping waves. At least the monstrosity
is exposed to all the vagaries of wind and tide; one can hope such insidious influences will
undermine it even if human planning doesn't eliminate it.
I can see it's time to think great thoughts again. I need a version of the Grand March from Aida
to deal with visual irritations. Perhaps imagining Russian dolls, one inside another, ad infinitim,
would be sufficiently compelling. Or perhaps the little person inside the head, he who is viewing
the vision seen by the eyes?
VISUAL CORTEX probably isn't the center for our perception of the faces and cars and trees
that we readily recognize. The utility-pole schema certainly doesn't live there, because none of
the cells in this region of brain can handle objects much larger than 2° (the sun and the
moon are both about 0.5°, just for reference).
The so-called visual cortex may get the information first from the thalamus, but there are many
other "secondary" visual areas in the back half of your brain. And they sometimes seem to do
fancier things. Perhaps triangles can be found there, and even the shapes of swans. Or my
schema for tumor-infested utility poles.
V4 is a patch of cortex where a complete map of the opposite half of the visual world was found
(as you might guess, V4 was the fourth one discovered in cortex). The left hemisphere's V4 has a
map of the right half of your visual world; the center of the retina is on one side of the V4
"patch" and the periphery on the other, the cells in between specializing in the intermediate
locations. The map is similar to one of those telethons where the people answering the calls from
the various states are stationed at desks that are spread out over the floor of a basketball court in
the shape of the United States, the "Maine desk" up in one corner, the "Texas desk" at the
bottom, etc.
In Daniel Hartline's metaphor, each brain cell is like a general getting verbal reports from the
battlefield that he cannot see directly. And so a brain map resembles a command center with
specialists in each battle sector. I remember those movies about World War II where England's
air defenses were coordinated from England-shaped plotting tables, different officers specializing
in each sector.
The receptive fields of V4 cells are often very similar to those found in V1. So why is there this
near-duplicate of V1? Is it like those backup command posts, there in case the main command
post was bombed? Of course, V4 isn't the only extra center with a complete map: In monkeys,
we've discovered several dozen so far. There are unlikely to be fewer in humans, and I wouldn't
be surprised if we had hundreds. That's why I call each half-map a "patch": The whole
convoluted cortical surface looks like a patchwork quilt when unfolded and flattened out, each
patch having a texture within it representing the fine-grain "grid."
Dozens or hundreds of "duplicates"? That's a lot of backups, so I doubt they are in fact
redundant spares in the manner of the airplane's three systems for lowering the landing gear.
Even if you argue that we have paired kidneys to insure against loss of one, then why not a spare
heart? For one thing, evolution by natural selection shapes up extras very slowly, as the Nth
spare would come under natural selection only on those rare occasions where all N-1 had failed
simultaneously. Evolution is full of good-enough solutions, where makeshifts are never
replaced; it seldom gets around to pair-and-a-spare.
But redundancy isn't just for backups: it can be a way of ganging up on a problem, as when a
dozen people get together to push a car out of a ditch. Having lots of separate centers doing the
same job can sometimes solve a precision problem, thanks to the Law of Large Numbers.
CONSIDER DUNKING FREE THROWS in basketball: You've got to be able to estimate how
far away that hoop is, if you are to drop the basketball through it cleanly. The distance is about
five paces, and the room to spare inside the loop is only a few fingers'-width -- so that's about
one percent of the total. While I doubt that basketball success played much of a role in hominid
evolution, hunters had the same problem when throwing at a small animal: about half your
chances of hitting a rabbit with a thrown stone occur hitting its front profile, but about half
involve the stone landing atop the animal. Hunters with good depth judgment will be twice as
successful as those with crude judgment. So how do I tell that the 20-cm rabbit is between 8.0
and 8.2 meters away, and not between 8.2 and 8.4 instead? That's again a judgment with a
tolerable error of about one percent.
We humans have a number of ways of estimating distance. Up close, we can see how much we
have to accommodate our eyes (squeezing that lens to bring the object into focus). Farther away,
the relative size gives one clue, as we know about how big adult rabbits are -- but you could be
fooled by an immature rabbit closer than you think. Texture is another clue: If we can see the
ruffles in its fur, it is closer than if we cannot. But the best method for distances under about ten
meters is the range-finder effect: We converge the two eyes to look at a close object, but make
their axes parallel to look at a very distant object. And some V1, V2, and V4 cells are very good
at making use of this.
In both V1 and V4, most cells receive inputs from both eyes. And it is a very similarly-organized
input: A receptive field mapped with the left eye shut is shaped the same as the receptive field
obtained looking only through the left eye. Looking with both eyes together usually gives the
most vigorous response. Now in V1, the best responses occur when the axes of the two eyes are
parallel, staring off at a distant object. But in V4, the two eyes' receptive fields are slightly
staggered, their centers not quite in the same place. And so the way to get an optimal response
out of the cell is to converge the two eyes slightly until the centers overlap perfectly. Thus the
cell, while responding to objects at various distances, responds best to those in a certain range of
distances, say between 7 and 9 meters away. Another V4 cell might prefer 2 to 3 meters because
its centers are even more offset and thus require more convergence to overlap. But V4 cells are
never sharply tuned, say, for 8.0 to 8.2 meters only and no other distance.
Should you average together the responses of a great many V4 cells, you can get a much better
estimate of distance. And usually these matters improve with the square of the number of cells:
With 25 times as many cells, you can improve your estimate by a factor of 5; a hundredfold
redundancy usually gives you a tenfold reduction in uncertainty. While the Law of Large
Numbers places some restrictions on the types of neural circuits that will provide this square-root-law improvement (the noise in the various cells must be statistically independent, each cell
must make a small contribution to the total so that they function as democracies rather than
oligarchies, the summation must be fairly linear rather than some binary logic, etc.), many types
of neural circuits can succeed in using the Law of Large Numbers to solve precision problems.
While I have worked it out in detail only for the case of differential depth discrimination and the
case of timing precision, it would appear that many types of precision discrimination behaviors
could profit from many-fold redundancy: Precision color discrimination (for telling ripening
fruit from the surrounding leaves), precision spatial discrimination (many animals exhibit
"hyperacuity" in a sensory system, where an individual's performance is better that the best
receptor's resolution), "perfect pitch," and so forth.
And so the prospect of several dozen extra visual maps immediately suggests a five-fold
improvement in some aspect of resolution, not several dozen backups in case of frequent failures.
It can be sensory performance that is improved, or it can be movement skills such as throwing.
Those basketball stars who always sink their long shots have probably learned to get a lot of their
brain cells to gang up on the depth discrimination problem and another group to gang up on
releasing their grip on the ball at just the right instant.
BUT THE SECONDARY VISUAL AREAS also differ from V1 when you look at optimal
stimuli. Often the differences are subtle, as when some V4 cells turn out to like different degrees
of convergence of the two eyes. Other V4 cells are much better at colored lights than are V1
cells. In some distant secondary areas such as the medial-temporal (MT) area, receptive fields
are organized strikingly differently from those of V1 or V4 cells; an MT cell will prefer dumbbell
or four-leaf-clover shapes rather than lines or corners. And so combinations of such elements
can come to specialize in irregular shapes such as the outline of a hand or the shape of a face.
Indeed, some such temporal lobe cells are said to like the shapes of faces better than anything
else -- maybe not exclusively your grandmother's face, but at least faces in general. So does one
have several dozen specialized subcenters?
Still, they overlap more than they differ, just what you'd expect if most had evolved by a simple
duplication: the same genetic instructions used a second time to make an extra map, with some
subtle modifications made later to subserve an additional function such as color or depth
discrimination. Duplication-then-diversification is a big principle at the gene level, a handy way
of improving a program while still running the reliable version a while longer, just as a computer
programmer always tries out modifications on a duplicate of the functioning program. It seems
likely that cortical map duplications were promoted by their usefulness when ganged together (on
occasions when real precision was needed), but that the duplicates later diversified a little and so
became useful for another function.
MORE THAN A QUARTER-CENTURY after Hubel and Wiesel found those line-orientation
specialists in visual cortex, no one has yet found a generalized triangle detector in any species'
brain. Nor are there many primate brain cells specializing in any of those features that the frog's
eye seemed to find so compelling in 1959. Why?
Well, perhaps someone will eventually find a generalized triangle detector cell, but we humans
remember so many different facts from our lifetimes that there may not be enough cells in our
brains for each cell to be a specialist in just one fact; just because computer memories store each
fact in a pigeonhole of its own doesn't mean that there aren't other ways to accomplish memory
and recognition tasks. Just because I can remember the profile of my grandmother's face doesn't
necessarily mean that I have a cell somewhere in my brain that responds only to that shape and
no other. Registering "that's a picture of my grandmother" could instead be accomplished by a
committee of cells, each of which also belonged to other committees (such as the one for my
father's face and for four-leaf clovers).
Expecting a specialist cell (or "labeled line") for each schema (Marvin Minsky, take note) is
called the Grandmother's face cell fallacy by neurophysiologists. It isn't that we're sure that such
a specialist cell doesn't exist (we haven't examined every brain cell yet!) -- it's that we are sure
that some properties emerge from combinations of simpler parts, that the whole can be created by
a sum of the parts. The parts may just be line specialists, and the whole created from lots of little
segments.
GESTALT PSYCHOLOGY emphasized, early in this century, that all experiences consist of
gestalten, integrated structures or patterns that must be apprehended as wholes rather than as
their disconnected parts. The notion that the whole is sometimes more than the sum of its parts
goes back to Aristotle -- but if it's not going to be just another leap of faith (what scientists call
hand-waving), if we are to understand more when we finish the explanation than when we started
contemplating the parts, then we need to surmount a difficult conceptual hurdle. Surprisingly, it
is closely related to another puzzle: how a function like perception and cognition need not have a
location, a place to which you can point and say, "There, Grandmother's face lives right there."
Happily, there is an excellent example of awareness based on committees and reducible no
further. This example has been learned (and then usually forgotten) by every neurobiologist,
probably in grade school. It is the lesson of color mixing, the first "emergent property" that we
came to appreciate. An emergent is a more-than-the-sum-of-the-parts property that emerges from
the collection of parts, and is incapable of meaningful reduction.
This sterling example dates back to 1802. It was discovered by the English scientist Thomas
Young, whose descendant John Zachary Young is one of the foremost neurobiologists of today.
J.Z. is well known around here, having discovered in 1936 the squid's giant axon (which is what
attracts so many scientists to MBL during the early summer's "squid season"). He went on to
analyze memory mechanisms of the octopus, write some of the major textbooks (you'll see The
Life of Vertebrates on many an MBL desk), give some of the earliest popular lectures on
neurobiology (the 1950 Reith lectures on the BBC), and write influential critiques of
neurobiology (such as Programs of the Brain).
Thomas Young discovered one of the great "you can't reduce it any further" themes of
neurobiology which stands as a warning to reductionists: just as the Nobska lighthouse warns
"you can't go any further and still float," so Thomas Young's analysis of color mixing now serves
to warn that you can't go any further than sensor committees without changing the agenda. Here,
at least, reductionism isn't everything: Committees are the real thing, at least if color is your
agenda. Or taste, for whether something tastes salty, bitter, acid, or sweet seems to be a matter of
irreducible combinations of activity in the chemical sensors embedded in the tongue; there are no
labeled-line specialists in one taste or another in your tongue.
Thomas Young got his start on this with the realization that we can see many different colors
(say, a hundred for the sake of argument) at each point (say, one billion) in our visual field.
Young suspected that the retina didn't have a hundred different sensors at each point in the visual
field, each labeled with a hue; he theorized that there were instead only three types of sensors at
each point and that it was the differing combinations of activity in each of the three that
communicated color sensations. A century and a half later, it was shown that he was exactly
right: We now call them the blue cone, the green cone, and the yellow cone (those colors being
the peak sensitivity of each photoreceptor type, not its exclusive specialty). Hermann von
Helmholtz in 1860 suggested that for each colorful stimulus, there is a ratio of responses across
these three sensor types that is specific to that color -- and that suffices to represent it. Right
again.
THAT A PARTICULAR SHADE OF RED is simply a 3:1 ratio in the activity of the yellow and
green cone types, with the blue cone mostly inactive, nicely accounts for the many different ways
we can produce the sensation of that reddish hue, one of which is with a pure 600 nanometer
wavelength. But that a single wavelength is usually the simplest way of producing the sensation
is misleading: some hues, such as purple, cannot be produced by any single wavelength.
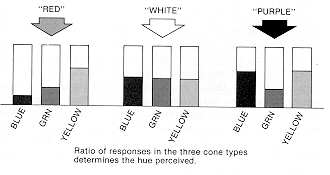
Purple corresponds to a pattern of activity in the yellow and blue cones (with green mostly
inactive) that cannot be achieved by any single wavelength, that indeed can only be evoked by a
combination of long and short wavelengths that would individually appear red and blue. Violet,
in comparison, is simply a lot of activity in the blue cone pathway, less in the green cone
pathway, and very little in the yellow cone pathway; it can usually be mimicked by a single short
wavelength between "blue" and "ultraviolet," such as 400 nanometers.
Evolutionarily, we should not expect pure wavelength specialists anyway: color is used to
distinguish one surface from another and, except for monochromatic gratings, all surfaces reflect
a combination of wavelengths. My proposed Purple Principle says that the need is for detecting
combinations (of which the two extreme ends of the spectrum together, but little of the middle, is
merely a rare example), not for detecting part of a rainbow (which is about the only time in
nature that we come close to seeing a single wavelength in isolation).
Of course, one can and should play reductionist games and analyze the component parts;
knowing that there are indeed three different kinds of cones, each with a different pigment and
thus wavelength sensitivity, is invaluable information. So is the fact that certain retinal ganglion
cells and geniculate cells specialize in color contrast. Some of these so-called "P-cells" might
even turn out to be far more sensitive to purple than anything else, and thus qualify as purple
specialists. But the existence of such narrowly tuned cells will not obviate the fact that color is
an emergent property of a committee of photoreceptors, capable of being expressed as the activity
of many broadly tuned cells as well as a few narrowly tuned cells. They serve to drive home the
fact that color hue is fundamentally a pattern and not a pure specialty with its own cubbyhole
somewhere in the brain whose activity signals purple to the mind.
What is so surprising is that, as Robert Erickson points out, neuroscientists are always
"rediscovering the wheel" while searching for labeled lines: The people working on skin
sensation rediscovered Young's principle, so did the people working on motor cortex's control of
movement. Young's patterning principle has been called many things by its reinventors:
population codes, parallel processing, distributed functions, ensemble coding, and across-fiber
pattern. It bears a strong resemblance to the holistic side of the reductionism versus holism
dichotomy. And to the developing distinction between the categories of cognitive psychology
and the distributed networks of computational neuroscience (to be discussed in Chapter 10).
Neurons are the anatomical units of the nervous system, but are not the
structural elements of its functioning. [The latter] have not yet been defined, and
it will probably be apparent when they are that they must be expressed in terms of
invariants of relative activities between neurons... and not in terms of separate
anatomical entities.... Unless [the observer] explicitly or implicitly provides a
theory that embodies the relational structure of the system, and conceptually
supercedes his
description
of the components, he can never understand it.
the Chilean neurophysiologist Humberto Maturana, 1980
The power of Young's logic is that the pattern is the message; it is the brain's
sensory code in its final form. This means that the population of neurons can
never be divided into separate neurons for, say, each discriminate color at each
point in the visual field.
the American sensory physiologist Robert P. Erickson, 1984
THE MOST FUNDAMENTAL REASON why labeled lines (another name for Grandmother's
face cells) aren't needed in sensory processing, however, has just occurred to me (though, before
fans of alliteration start calling this Calvin's Coactivated Committee Concept, we'll have to see
how many other people also recognized this minimalist principle during the last century). We
don't need a single cell specializing in "purple" simply because it takes many cells for me to
pronounce "purple" (or act on the purple-coded information in other ways). Many-to-many will
suffice without a many-to-one-to-many hourglass-shaped bottleneck.
Sensory processing is all about coupling sensation to action in a manner that is reliable. And
actions all require a motor program that orchestrates a number of muscles, creates a
spatiotemporal pattern of cell firing something like a fireworks finale. An action always involves
many muscles -- and therefore many motorneurons. This usually involves some motorneurons
firing first and others later.
Triggering this ensemble into action need not require a single "command neuron"; it only
requires a committee of interneurons active in some characteristic pattern. It is the difference
between starting an automobile engine by pushing the Model A's old-fashioned starter button and
the modern procedure: Turning the correct key in the ignition lock where each tumbler has to be
set just right. It is the correct notching pattern that starts the engine of a modern car, not a single
push on a single button.
The analogy is deficient in that it is the activity in a single information channel (the wire from
ignition switch to starter motor) that really starts the car. Manufacturers who want to slow down
thieves use a ribbon cable from switch to effector, fixing it so that a pattern of information in
many wires is required rather than touching a single wire to the battery connection. A keypad
outside a door, connected to the electrical door lock, needs to use a ribbon cable to foil thieves
who merely rip the keypad from the wall and touch two wires together.
Animals seldom use the single-wire approach; the patterned activity in a ribbon cable is the more
appropriate analogy for almost all motor programs. The classic exception is the Mauthner cell in
the brain stem of fish: When it fires a single impulse, the fish executes a massive tail flip. One
could put a label on the right side's M-cell saying "emergency on right, flip left" and another on
the left M cell saying "emergency on left, flip right" and have true labeled lines.
It is only very simple motor programs with simple spatiotemporal patterns of muscle activity that
can get by with the "Model A" approach to orchestration. The appropriate trigger for most motor
programs is going to be a keylike correct combination of triggers in many cells; indeed, it will
probably be just as important which cells are inactive as which are active. Therefore, one
expects the ultimate stage in sensory processing to produce a pattern as the trigger. And it's not
just a spatial pattern like the key notches: it is a spatiotemporal pattern, like the fireworks finale,
the order in which various neurons are activated, as well as which neurons are activated, being
the key.
The sensation-to-movement transformation is many-to-many; there is no need for a many-to-one-to-many bottleneck unless the one cell has some special advantage for producing the spatial or
temporal aspects of the movement subcommands (as does the M-cell). That's why purple doesn't
require a specialist neuron: The correct combination of activity in yellow cone and blue cone
channels (together with the absence of green cone channel activity) should suffice to pronounce
"purple." That's why labeled lines and command neurons are so rare, why distributed sensitivity
and committees are so common.
To categorize is human,
to distribute, divine.
Terrence Sejnowski, 1988
PATTERNS AS THE DETERMINANT, rather than absolute quantities of some one thing, are
also a big feature of growth and development. The curling vines that one sees along the bike trail
are good examples of this.
How does nature produce a nice, graceful, regular curve? Or a spiral? Largely by making one
side of the stalk grow faster than the other. Want to make a stalk bend to the south? Just let the
north side grow faster than the south side. Want it to grow in a corkscrew? Let the east side at
the base also grow faster than the west side. Want it to spiral upward, tightly coiled like a
telephone cord? Just keep the upward growth rate low compared to the north-south and east-west differences. The final shape is "specified" by the relative magnitude of those rates, not by
individual magnitudes.
Forms can result from such simple rules involving differences and ratios in growth rates. The
genes do not contain an image of a spiral: They contain a set of codes for enzymes. Those
enzymes control growth rates. One pattern of enzyme quantities results in a tightly curled spiral,
another pattern in a stem that merely tilts south. It is another example of the Purple Principle.
The curved surfaces of our bodies are due to such differential growth rates too, a surface layer of
cells dividing faster than a deeper layer of cells and so bending the sheet. The shape of a dog's
face, whether pointed like a setter or flat like a pug, is a matter of differential developmental rates
too: Puppies tend to start off flat-faced, and some grow the elongated noses of setters while
others have their growth terminated before ever getting to that stage. When the development of
sexual organs proceeds faster than general somatic development, the body form often gets
arrested in a juvenile form, since puberty tends to slow down further somatic development to a
crawl. Animal breeders have, in effect, selected for those variants with markedly different sexual
versus somatic developmental rates in breeding for the pug.
And so it makes remarkably little sense to ask "where" the stem's shape is stored in the plants'
genes or where the face's shape is stored in a mammalian chromosome. It's going to be a pattern,
like purple.
CLOSELY RELATED to the Grandmother's face cell fallacy is the little-person-inside-the-head
fallacy, the notion that there is some ultimate emperor to whom all those specialized command
centers report. It's reminscent of those Russian dolls that come apart, only to reveal a slightly
smaller version of the doll inside. It too comes apart.... Now at some level, there will be too few
atoms left for the shape of a doll to be maintained; you can keep taking it apart into protons and
neutrons and electrons, and then into all their quark components -- and that is proving very
interesting -- but you're no longer talking about miniature dolls. You've changed the agenda.
I've long been suspicious of an infinite regress: It always reminds me of something that
happened when I was about six years old. One snowy Saturday morning in winter, my father
took me down to the local barber shop for a haircut. I'd always liked the barber shop, which was
full of new smells and absorbing rituals. In those days, it was something of an all-male club, at
least on Saturdays, when fathers brought their sons in. It was a father-and-son sort of thing,
something like following the fire trucks to see a fire (my father was an executive of a fire
insurance company, and so I learned how to tell the sirens of the fire trucks from those of the
police cars from those of mere ambulances; for my birthday each year, I'd get to go to the local
fire station and ring the brass bell on an old fire truck, once for each year of age).
The barber shop was long and narrow: You saw a long row of barber chairs down one side of the
room, with waiting chairs and magazines down the opposite side, the shorn and the unshorn
facing one another. The shorn towered over the unshorn. Young boys sat especially high up in
the chair, boosted up by a special board placed across the arms of the old-fashioned chairs so that
the barber didn't have to bend over. And so you had this commanding view over everyone's
heads: and the view was overwhelmingly of yourself.
Down each wall were mirrors, big plate glass mirrors. No matter which way the chair rotated,
there was the mirror reflecting back your image -- as well as the image of the mirror in back of
you showing you what you looked like from the rear. But of course it too contained a somewhat
reduced-size image of your front. Since someone had carefully made the mirrors parallel to each
other, there was an infinite regress of ever-smaller images of your front, back, front, back, front,
back.... I had marveled at this Russian-doll-like succession during my first visit to the barber
shop, but later tired of it.
Then came one quiet Saturday of winter when it was softly snowing outdoors. I was perched
atop the chair, getting bored. There was some commotion outdoors, but I couldn't see anything
because I was a few chairs down the long corridor from the front window, not in the first chair as
I liked to be. Various people disappeared to see what was going on, and I became even more
impatient for the barber to finish. A siren or two was heard, but then nothing. An accident of
some sort, according to the people (including my father) who'd looked out the window. And
there I was, trapped in this glorified high chair.
This haircut seemed to take forever. The barber tried to get me to count my images in the mirror,
saying that there was a prize for the boy with the sharpest eyes who could spot the smallest image
in the center of it all and see if the smallest one was different. And so I diligently counted this
infinite regress, but they all looked the same to me: None was different -- all were me -- except
for being a little smaller each time. So the barber told me to count them again.
There was much fussing with talcum powder, even some fancy-smelling hair tonic that I'd never
had before. Finally, I was unwrapped and helped down off of my high perch. My father paid the
barber, thanking him more than usual. Then I was wrapped up again, this time because of the
winter cold. Eventually we went outside into the fresh snow. I'd almost forgotten about the
earlier commotion.
The barbershop was near the end of the trolley line. A big circular loop track allowed the cars to
turn around and go back into the city; I had often watched the trolley cars clanging their way
around this circle, warning the pedestrians who took shortcuts across the tracks. Now, a cluster
of people gathered around an abandoned trolley car, and a crumpled figure lay on the ground
covered by a blanket. A smaller object lay a few feet away, covered by someone's coat. His leg.
The dark reddish-brown stains in the snow were not at all like the catsup-red of blood in the
make-believe of movies. All was being slowly covered by fresh snowfall. It was, the people
said, a sad case of suicide.
Each time that I encounter another infinite regress, I wonder whether it is just another time-consuming exercise. And if it will lead to another dead end, literally at the end of the line.
[If] a command to move originates in prefrontal cortex, that command should be
considered a product of multiple interactions of prefrontal cortex with other
cerebral components, cortical and subcortical. Thus the quest for a prefrontal
executive is pointless. Only by reasoning this way do we avoid an infinite regress
of ever higher executives or the implausible notion of prefrontal cortex in a
pontifical position.
the American neurophysiologist Joaquin Fuster, 1981
PHILOSOPHERS TRADITIONALLY COMPLAIN about infinite regresses and tautologies, but
practical people like electrical engineers aren't anywhere as worried. My old friend John DuBois
points out that most oscillator circuits tend to chase their tails; you just have to use the right types
of phase plane reasoning in order to analyze such feedback circuits (as an undergraduate, he tried
pointing this out to his philosophy professor but got nowhere). Cause-and-effect reasoning
sometimes isn't very good when it comes to open systems with energy to spare. Might a semi-infinite regress subserve consciousness, some sort of frontal lobe circuit that chases its tail?
We like to ridicule bureaucracies whose committees do nothing but "shuffle papers," making
endless recommendations to one another but never acting. "Paper chasing paper in a circle." Yet
the cautious bureaucracy may be a good analogy for our subconscious. The problem is: At what
point does one act, do something in the external world? What's a good analogy to decision-making?
Homunculi are only bogeymen if they duplicate entirely the talents they are rung
in to explain.... If one can get a team of relatively ignorant, narrow-minded, blind
homunculi to produce the intelligent behaviour of the whole, this is progress.
the American philosopher Daniel C. Dennett, 1978
Email ||
Home Page || Table of Contents || Endnotes for this chapter || continue reading Next Chapter

