 |
A book by William H. Calvin UNIVERSITY OF WASHINGTON SEATTLE, WASHINGTON 98195-1800 USA |
|
THE CEREBRAL CODE Thinking a Thought in the Mosaics of the Mind Available from MIT Press copyright 1996 by William H. Calvin |
|
Convergence Zones with a Hint of Sex
Long-distance communication within the brain has to bear the task of matching up information across sensory modalities. I say that because, though there are multisensory neurons in the cortex, the stroke evidence suggests that the memory traces for the visual aspects of an object are kept pretty close to visual cortex, while that for auditory aspects of the same object are kept on the periphery of auditory cortex. Yet we associate them pretty well, most of the time. Here’s what I said on the subject in How Brains Think:
So how do we integrate those separately-stored codes into a master code for comb? Or its equivalent, a process somehow able to associate all those aspects with one another?
|
|
Distances in the brain are enormous, compared to the scale of millimeter-long dendrites. My
longest uninterrupted axon is probably the one that runs as a thinner-than-hair thread from the tip
of my big toe to the dorsal column nuclei at the top of my neck, the better part of 2,000 mm. Most
of the business of deciding to send an impulse off on that long trip is done within a 0.1 mm segment
at the beginning of the axon. It also takes a while to make the trip, and measuring distance in terms of travel time often makes more sense than thinking in terms of miles or kilometers. There are similar considerations in the brain: I suggested earlier (p. 36) that my triangular arrays might be better measured in travel time than in actual distance. Axon conduction velocities vary enormously, even in different branches of the axon of one neuron. Sending an impulse from one hemisphere to another takes as long as sending it to the spinal cord, even though there is a tenfold difference in distance. That’s because the corpus callosum path is so slow. Faster conduction would require more myelin wrapping, so fewer axons would fit in the limited space of the corpus callosum. In the monkey, about 70 percent of the axons in the corpus callosum at birth are withdrawn during the first six months of postnatal development. I say withdrawn because neuron death in that period is fairly low; the 70 percent figure surely represents the withdrawal or pruning of some of the branches of a neuron’s axon, not the death of the entire treelike neuron.
|
 And what neurons are these, anyway,
that send their axons over such long distances between hemispheres and front to back within a hemisphere? The major
pipelines for long corticocorticals, such as the arcuate fasciculus
connecting temporal and frontal lobes, have long been noted. But
there are also spotty projections, discovered a half-century ago in
the days of strychnine neuronography. They are quite widespread
and modern neuroanatomical techniques have revealed that they
may be organized into macrocolumns and layers. So what are the
pipeline and spotty long-distance projections doing? Surely they are biasing the distant basins of
attraction, at a minimum. But might they clone hexagons like a <
And what neurons are these, anyway,
that send their axons over such long distances between hemispheres and front to back within a hemisphere? The major
pipelines for long corticocorticals, such as the arcuate fasciculus
connecting temporal and frontal lobes, have long been noted. But
there are also spotty projections, discovered a half-century ago in
the days of strychnine neuronography. They are quite widespread
and modern neuroanatomical techniques have revealed that they
may be organized into macrocolumns and layers. So what are the
pipeline and spotty long-distance projections doing? Surely they are biasing the distant basins of
attraction, at a minimum. But might they clone hexagons like a <|
There are two types of corticocortical projections, those that stay within the cortical layers and
those that loop down through the white matter. The former, which are the intrinsic horizontal
connections of the earlier chapters, are mostly local (though that can mean a few millimeters). The
latter can go long distances, as from one hemisphere to another through the corpus callosum,
though most only make a U-shaped passage through the white matter of one gyrus and then
terminate in a nonadjacent patch of cortex that’s only a few centimeters away. All of those axons can come from one of the superficial pyramidal neurons we have been discussing. The long axons taking the white-matter routes typically terminate (though not exclusively) in the same superficial layers, once they arrive in distant cortex. So far, we’ve been looking at the ramifications of their axon branches that run sideways within layers 2 and 3, without ever looping through the white matter. These sidestepping connections are perhaps analogous to internal phone calls to neighboring offices, the U-axons to local calls within the city, and the lengthy U’s to long distance calls. Some cortical areas might be what Damasio calls convergence zones, a focal point for disparate modalities. I think of a convergence zone as something like the long-distance operator that sets up conference telephone calls, serving as the center of a funnel that rebroadcasts. Of course, the linkage could be centerless, as when a conference call of a half-dozen committee members is achieved by chaining, each person using their second phone line to link yet another member to the conversation via their phone’s conference button. The analogy to the telephone network may be misleading, both because of the phone system’s point-to-point nature and because of the way open connections are established and maintained until hangup. Packet-based networks demonstrate an alternative, as when web pages are displayed and manipulated on the client machine without further attention from the server or the intervening network. We tend to think of those long corticocortical axon bundles as if they were fiber optic bundles that convey an image by thousands of little light pipes.
And the fanout of connections at the far end (p. 19) is most unlike optical fiber terminations in another, more serious, way. A given axon fans out to connect with dozens of recipients, scattered over the better part of a millimeter. Such a point-to-area mapping, in the manner of a flashlight beam, only makes matters worse for any point-to-point mapping. So, at first glimpse, it appears that corticocortical bundles are considerably worse than those incoherent fiber optic bundles that are factory rejects — unless, of course, something else is going on, not captured by our technological analogies to fax machines and fiber optics. Indeed, triangular arrays give one a different perspective on the fanout “problem” — even converting it into a virtue.
|
 Of course, light
pipes can go astray if the bundle is not
carefully assembled; manufacturers have to
make sure that neighbors at the sending end
remain neighbors at the receiving end. An
incoherent fiber bundle can create displaced
facial features à la Picasso and neural wiring
surely has many such imperfections.
Of course, light
pipes can go astray if the bundle is not
carefully assembled; manufacturers have to
make sure that neighbors at the sending end
remain neighbors at the receiving end. An
incoherent fiber bundle can create displaced
facial features à la Picasso and neural wiring
surely has many such imperfections.
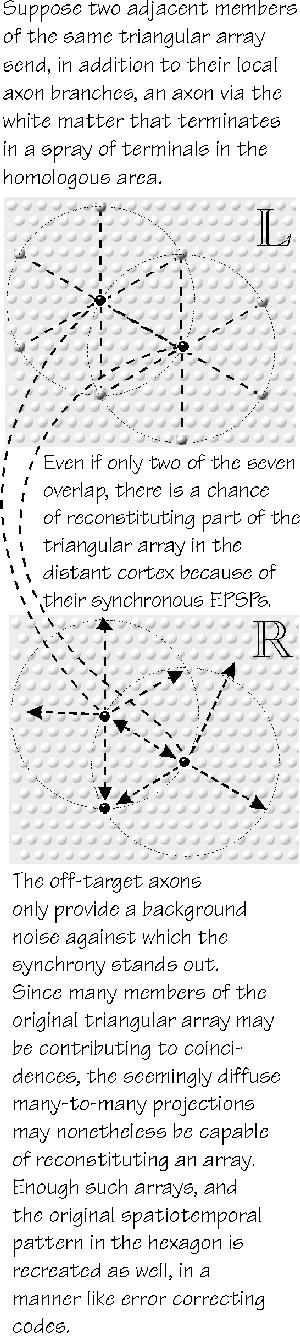 Hexagonal cloning with error-correction standardization could also
happen in the target cortex, reproducing the spatiotemporal pattern that is
being maintained on the sending end.
I don’t mean that in the sense of
video point-to-point mapping, but
rather in the sense of reconstituting the
“weave” of the fabric
without mimicking the extent of the
fabric swatch (that dynamically
changing patch on the quilt). In this
scheme, you don’t reconstitute
the whole choir but a lesser number of
singers, yet they’re singing the
same song and may themselves recruit
neighbors to form another choir.
Perhaps even a larger one.
Hexagonal cloning with error-correction standardization could also
happen in the target cortex, reproducing the spatiotemporal pattern that is
being maintained on the sending end.
I don’t mean that in the sense of
video point-to-point mapping, but
rather in the sense of reconstituting the
“weave” of the fabric
without mimicking the extent of the
fabric swatch (that dynamically
changing patch on the quilt). In this
scheme, you don’t reconstitute
the whole choir but a lesser number of
singers, yet they’re singing the
same song and may themselves recruit
neighbors to form another choir.
Perhaps even a larger one.We need to start with one note of that melody and its triangular array, indeed, with only one of its synchronized cells. There currently isn’t data on the distant terminal fanout of one axon, nothing comparable in detail to that for the point-to-annulus fanout of the close-in branches, with the silent gaps described in chapter 2. We know that distant-terminal fanout exists, spanning macrocolumnar dimensions, but not much more. So I had to make a theoretician’s assumption about the long-distance-call termination: that it is, on average, a point-to-annulus fanout, just like the one back home in the 0.5 mm around the parent neuron (or minicolumn, p. 42) that structures the short path sidestepping. It’s like a flashlight beam with a central bright spot, plus a bright peripheral ring. That little assumption buys quite a lot compared to the point-to-area alternative. Any one point in the target cortex is going to get inputs from a number of points in the sending cortex. How many? It might get one from the homologous point back home, directly on target for a point-to-point mapping. But there is also an active triangular array back home, all firing in synchrony. So the same point in target cortex might get another input from the fanout of axons from an adjacent point in the sending triangular array. Indeed, six such triangular nodes back home could make contact with the distant point in this manner, for a total of seven synchronous inputs — if nothing is lost along the way.
Let us say that half the potential axon terminals are lost. Still, it may
only take a few synchronous inputs to recruit a neuron, given enough repetition. If the same thing
happens 0.5 mm away, the pair can get their own triangular array going. Temporal dispersion may
be less of a problem, as well. The synchrony criterion allows for a fair amount of time shift between
the two arrivals; temporal summation of inputs depends on the decay times of the PSPs, and a few
milliseconds dispersion might not matter very much. The terminals in the superficial layers suggest
that NMDA mechanisms are involved and, given all the repetition to clean out the Mg Even allowing for a fair amount of imprecise topographic mapping and a certain amount of temporal dispersion along the way, I think that a large enough sending area can get a small territory of similar arrays going in the target cortex (except, perhaps, for a change in the “0.5 mm” metric to reflect that typical of the target cortex). Error correction can shape up a standard version of the one-note triangular array in the distant cortex. If that happens for many of the triangular arrays, the spatiotemporal pattern at the target cortex is likely to be close to the tune playing back home. The distant terminals might also prove to have the second annular ring of fanout branches as seen back home at twice the local metric distance; that would provide another six possible synchronized inputs. Only a few of the thirteen could suffice to start up a repeater node; only two such adjacent repeaters could suffice to start a local triangular array that can extend itself.
So, we have created a distant repeater of the triangular array back home. And that applies to each of the triangular arrays there, each interested in different features of an input (or contributing to different aspects of an output). We have cloned the hexagon in distant cortex, the faux fax at last. And it is just the type of mechanism needed for linking together distributed features of a database, the question raised in chapter 7.
|
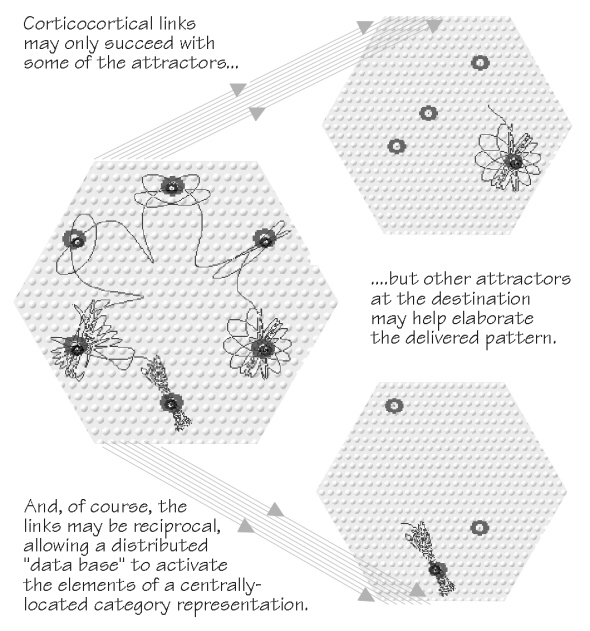 The strays will, of course, provide a certain amount of background
noise. They might fog the pattern (p. 107) but there are two ways of getting around that. First, just
as the view from two eyes separated by a few centimeters can disambiguate scenes that are too
noisy for one eye alone, so we are bringing to bear a half-dozen or more elements of the sending
array, all converging on a point. Second, I suspect the local
The strays will, of course, provide a certain amount of background
noise. They might fog the pattern (p. 107) but there are two ways of getting around that. First, just
as the view from two eyes separated by a few centimeters can disambiguate scenes that are too
noisy for one eye alone, so we are bringing to bear a half-dozen or more elements of the sending
array, all converging on a point. Second, I suspect the local |
Two adjacent hexagons, once started up in the distant cortex and singing the same music, are
sufficient to clone a choir producing the same melody, but success will depend on the exotic
(literally, in another place) resonances. This spatiotemporal pattern’s relative success in
cloning competitions ought to depend on the same types of factors as in the sending cortex: biases
to the local basins of attraction. But they ought to differ in detail, to arrive against a different set of resonances, biased by different ratio of neuromodulators — and so the competitive outcome could be very different than back home. Just as emigrants may thrive in distant places, so faux faxed spatiotemporal patterns might find a completely different reception in the distant cortex. Typically, of course, dying out without issue, but occasionally thriving. Not all of the parent attractors may arrive intact; even when they do, they arrive against a very different background of passive attractors than existed back home, both from short-term and long-term memories residing in the target hexagons. Furthermore, there may be several faux faxes arriving at the same time from different senders — which means we have a new form of competition, quite in addition to the side-by-side dog and cat pavers type of lateral competition for space introduced in chapter 4. For all these reasons, the spatiotemporal patterns in the target region may not be mere superpositions of the contributors, those jazz performance overlays. In Hebb’s other dual trace memory system, you can get melding of attractors in “bilingual belts” from noncontemporaneous occupations of the target area. Such exotic combinations are best introduced, probably to no one’s surprise, after a preliminary reminder about sex.
|
|
Among the accelerators of evolution is sex, both in the sense of recombination but also in
Darwin’s sense of sexual selection serving to create elaborate peacock tails much more
quickly than environmental selection would ever do. Among all our dichotomies, is there anything
that could be analogous to sex? The novel aspect of the biological invention of sex was not the exchange of genetic material (bacterial conjugation and retroviruses had likely been busy doing that for some time) but the development of more specialized vehicles called gametes that had storehouses of energy, handy for either mobility or fetal development. And the interesting thing about gametes is that equal-sized ones weren’t stable; once there were slight variants in stored metabolic energy, evolutionarily stable strategy considerations took them to the extremes of little sperm and big ova. This gamete dimorphism is what gives rise to most of the secondary characteristics of males and females, including their differing reproductive strategies. Because sperm are cheap (an adult human male may produce 40 million a day), one male has the potential of fathering an almost unlimited number of offspring. Ova are expensive and so one female has a more limited number of potential offspring (a human female is born with her lifetime supply of a few hundred, which is still an order of magnitude more than she can possibly rear). On the other hand, a female is almost guaranteed some offspring, at least in comparison to a male who may well get none at all because of the competition for access to females. In many species, this leads to females being choosy about sexual partners at the same time that males are more indiscriminate.
To ask if such kinds of sexual selection could be going on in a system such as neocortical hexagonal competitions, it is not necessary to identify a traditional male/female distinction. For example, a simple and a complex code, with a tendency to merge, might suffice. So long as there are at least two general types of codes, differing in some inheritable property that affects reproduction, one can have something like sexual selection operating.
One set of extremes was seen in the chapter 7 analogy to the hash versus the full text, where mere recognition can be done cheaply (hash the incoming and compare it with a low-dimensional database of hashes of the higher-dimensional stored memories), but recall is more demanding, requiring an expensive investment in reconstructing detail (probably starting with an intermediate representation, the prototype or some other form of loose-fitting abstract). Still, complex spatiotemporal patterns (up to the size that can still be contained in a hexagon) can clone, just as simple ones can. But the production of new individuals (as opposed to just another clone) can involve both error and recombination. Is there any possible sexlike dichotomy in recombination? We now have three types of superposition: those “in-house” ones associated with the intrinsic horizontals and borderline superpositions, the “local call” ones associated with U-fiber projections within a cortical area (and often within a particular sensory modality), and the “long distance” ones where different sensory and motor modalities may converge. We also have several superposition mechanisms, the active ephemeral ones and the new attractors in the connectivity that might blend experiences from different times and places in belts of overlap.
|
|
Superimposing two stanzas is perhaps more difficult than adding a simple trill to an existing
stanza. Superimposing a complex spatiotemporal pattern upon
another complex one, in such a way that it can be embedded in the
connectivity and later reconstituted, is surely a more difficult task
than successfully superimposing two simples or superimposing a
simple on a complex. In biology, there is always the issue of the recombination’s viability. Most are unsuccessful in reaching reproductive age, whether from juvenile mortality or (as naturally happens with 80 percent of human conceptions) spontaneous abortion. Matings between related species can occasionally produce hybrids, though many are dead ends because of sterility. So, too, we have compatibility issues when dealing with a hexagonal connectivity that simultaneously supports many different basins of attraction: some superimposed spatiotemporal patterns are going to be captured by the existing attractors, others are going to fail to be incorporated, but a few might successfully change some aspect of the hexagon’s repertoire (likely only a trills’ worth of one of its many melodies) via a connectivity alteration.
The recombination issue could be avoided, of course, if the connectivity were a tabula rasa: so plastic that the hexagonal mosaic became a buffer, holding only one very detailed attractor in its connectivity (and not dozens). Intermediate levels of plasticity might allow multiple melodies from the same connectivity, or they might allow anything more than a week old to be overwritten. But one can imagine a hexagon’s worth of connectivity that added attractor lobes only reluctantly, and only to some of its dozens of separate attractors. That would mean that simple-complex pairings would be the most common “new individuals,” loosely analogous to the way sexual reproduction forms new individuals by recombining a small and a large gamete. Tacking on an additional feature creates, of course, an extra-strange attractor (given that attractors with multiple lobes were originally called strange attractors, as in the two-lobe butterfly attractor). This makes our hint of sex in convergence zones extra strange.
|
 Chimeras, as in the she-monster of Greek mythology that had the
head of a lion, the body of a goat, and the tail of a snake, are considerably more rare than hybrids.
Still, occasionally an individual is seen with odd features such as two blood types that suggest
fraternal twin fetuses amalgamated into one individual early in gestation, or that a mother gained
some cells from a gestating fetus. One can imagine similar amalgamations of attractor basins, just
as in those Charles Ives melodies with an intruded snippet of Yankee Doodle. Though copied
everywhere, such an intrusion might have meaning in only one cortical area, just as only part of the
genome is decoded by the specialized cells of the liver but all are copied during mitosis there.
Chimeras, as in the she-monster of Greek mythology that had the
head of a lion, the body of a goat, and the tail of a snake, are considerably more rare than hybrids.
Still, occasionally an individual is seen with odd features such as two blood types that suggest
fraternal twin fetuses amalgamated into one individual early in gestation, or that a mother gained
some cells from a gestating fetus. One can imagine similar amalgamations of attractor basins, just
as in those Charles Ives melodies with an intruded snippet of Yankee Doodle. Though copied
everywhere, such an intrusion might have meaning in only one cortical area, just as only part of the
genome is decoded by the specialized cells of the liver but all are copied during mitosis there.|
But this new individual isn’t doubly strange: it doesn’t carry around nearly equal
numbers of contributions from both parents in the familiar heterozygous manner, with two alleles
often available. Think of the offspring as very much like one parent (indeed, it is that parent,
modified — but since the parent exists in numerous hexagonal clones, and only a few may
be modified, the parent pattern may live on elsewhere), with just a touch here and there of the other
parent.
What the new individual has, rather than all those alternative alleles
of the truly heterozygous, may be a working link to the minor parent: the umbilical cord
hasn’t been broken. That trill’s worth of addition could be the hash that, elsewhere
in neocortex under some circumstances, One can imagine reciprocal connections or strange attractors that work too well, as when a stimulus in one sensory modality evokes a strong memory in another (an element of the condition known as synesthesia). Links that fail could presumably give rise to a variety of minor (anomia) and major (agnosia) complaints, simply from the failure to make accustomed links. An attractor’s complexity may, or may not, be expensive to create (simple rules can have complex consequences), but it is surely expensive to maintain over a long haul, simply from what economists would call opportunity costs (in this case, lost opportunities, foregone options in the name of persistence or stability). No matter how large your hard disk, it rapidly fills up; surely your brain has the same accretion problem, even in childhood. Maintaining attractors has a cost, commonly noticed as lengthening access times that slow performance at some tasks (a subject to be addressed later, at p.185).
|
 evokes a full-text spatiotemporal pattern and sends it back.
The new individual could inherit from its major parent a whole collection of such links to outlying
hexagons and their more detailed attractors. So here we have not only a candidate for what integrates the sight-sound-feel-pronunciation of comb but a suggestion of how we might tack on a new
attribute (those broken teeth that help identify it as my comb!) with a trill-like link.
evokes a full-text spatiotemporal pattern and sends it back.
The new individual could inherit from its major parent a whole collection of such links to outlying
hexagons and their more detailed attractors. So here we have not only a candidate for what integrates the sight-sound-feel-pronunciation of comb but a suggestion of how we might tack on a new
attribute (those broken teeth that help identify it as my comb!) with a trill-like link.|
Concrete thinking provides one example of premature closure, of terminating a search for mental
links too soon. And we can now see one way in which this could happen: with a different plurality
for successful links than for successful closure. The simplest model for closure is the movement choice example of chapter 4 (p. 57), where various candidates for a hand movement competed until some plurality was reached, at which subcortical mechanisms launched the movement with the strongest chorus. Call this plurality requirement the N for action, Naction. Activating the kth link out of this same territory takes Nk to get the same spatiotemporal pattern going with smaller numbers in the target cortex. Suppose closure for action normally takes a chorus of 100 hexagons, while establishing a link only takes 50. Then suppose that Naction is lowered to 40 without changing the linkage requirements, or that the Nk requirements were raised to 125 because of signal/noise ratio problems at the destination. In either case, you would act without considering some of the linked attractors stored elsewhere. And so you would have trouble seeing analogies. In extreme cases, you might even dissociate the various sensory representations of the same object, as in the agnosias. The half-second-and-longer reaction times seen in much cognitive processing have always been a puzzle from the standpoint of conduction times and synaptic delays, all an order of magnitude briefer. Add to the time to recruit a local chorus the time needed to form a second territory via a link, and cross-modality matching experiments might require the better part of a second, simply because of the repetitions needed to gradually develop the links. With a darwinian process operating in cerebral cortex, you can now imagine how stratified stability could generate a strata of concepts that are inexpressible, except by roundabout, inadequate means — as when we know things of which we cannot speak. Decomposing them via successive links into speakable concepts is a lot of additional work beyond that point at which you sense the problem’s criteria have been fulfilled.
|
|
Corticocortical improvements are an interesting issue here, as surely the corticocortical axons
do something simpler most of the time, not arbitrary spatiotemporal patterns. The axon fanout at the destination, if not tuned up for recreating triangular arrays, presumably produces a distorted version of the spatiotemporal pattern at the origin. Presumably the receiving cortex handles this in the manner of categorical perception, tuning up to recognize special cases. When the number of vocabulary items is dozens to hundreds, this probably suffices. We, however, have vocabularies of 105 and can extend them to novel concepts, such as when we talk about how many angels can dance on the head of a pin. That suggests we have improved our corticocorticals to the point that they can transmit arbitrary spatiotemporal patterns. I will discuss this again at the end of the final chapter, when I engage in a brief digression into Universal Grammar, but here I wish to point out the route to improving corticocorticals from special-case codes to arbitrary codes.
Assuming a Darwin Machine already exists on both ends, then the size of the sending array could be a major determinant of whether the origin’s spatiotemporal pattern could be recreated at the destination.
|
|
Long-distance connections, one is tempted to claim, are something like an extended family. If
an area’s local history of cloning competitions serves to create a community of interacting
individuals, then the long-distance links are something like the scientific community in the early
days of the Royal Society, that interacted mostly by letters with many copies, and later, publications. Or like the virtual communities of cyberspace. Such interacting groups don’t have all the features of local communities — you can’t, for instance, borrow a cup of sugar from the neighbors. But then you don’t have to listen to the neighbor’s barking dog, either.
|