 click for enlargement of this Mark Meyer painting |
THE CEREBRAL CODE Thinking a Thought in the Mosaics of the Mind Available from MIT Press copyright ©1996 by William H. Calvin |
1. There is a characteristic pattern involved. We have identified a pattern (spatiotemporal firing pattern within a "0.5 mm" hexagon in the superficial layers of neocortex) that can be standardized in a crystalline manner (it's Dawkins' minimum replicable unit). The pattern is as abstract as a bar code; just as the DNA string looks nothing like a folded protein, these cerebral codes bear little resemblance to the objects or movements they represent. 2. The pattern must be copied somehow (indeed, that which is copied may serve to define the pattern). Copying identified a way of compacting the essential Hebbian cell-assembly, and its connectivity changes, into only two adjacent 0.5 mm hexagons of neocortex. Thanks to the point-to-annulus tendencies of the same neurons that recurrently excite one another, two adjacent "parent" neurons seem capable of entraining a third and fourth to initiate a triangular array, one that can be extended for some distance. But the overall effect is to clone the spatiotemporal pattern of all the active triangular arrays within a hexagonal-shaped area; thus we can often speak of hexagonal cloning. Even noncloning hexagons may have an important function: to serve as a barrier. 3. Variant patterns must sometimes be produced by chance. Variants can be generated by relaxing error correction at gateways through the barriers. But superpositions readily occur; because these patterns are a fairly sparse fill of the hexagon, the superpositions may be able to code for supersets such as categories, sensory-motor associations, and even relations between relations such as syntax and metaphor. It all doesn't have to be crammed into one hexagon because "message digest" hashes may suffice to call forth the "full text" from elsewhere. 4. The pattern and its variants must compete with one another for occupation of a limited work space. Indeed, with end runs around barriers, competition is hard to avoid. The cortex is certainly limited, just as is a patchwork quilt. The work space is probably the equivalent of that paved park, with its alcoves in which demes can go extinct. Whether all neocortex functions this way or whether specialization may often prevent cloning competitions is an empirical question to be answered as experimental resolution improves. 5. The competition is biased by a multifaceted environment, for example, how often the grass is watered, cut, fertilized, and frozen, giving one pattern more of the work space than another. No, we haven't yet identified which cortical pattern is the crabgrass. But long-term memory is a memorized environment (via connectivity changes that create basins of attraction). Sensory inputs from our real-time physical environment (and more distant cortical inputs) should be able to bias resonances without actually cloning distinctive spatiotemporal patterns. Short-term memories in the form of fading basins of attraction, established by cloned patterns a few minutes earlier, can form part of this environment and so facilitate stage-setting moves. 6. There is a skewed survival to reproductive maturity (environmental selection is mostly juvenile mortality) or a skewed distribution of those adults who successfully mate (sexual selection), so new variants always preferentially occur around the more successful of the current patterns. Nothing I've said so far has demonstrated that the more populous patterns have a tendency to generate most of the next generation's variants -- but perhaps you've guessed why. It's the ubiquitous surface-to-volume ratio relationship that provides Darwin's inheritance principle. In a basically two-dimensional world such as retina or cortex, it's called the perimeter-to-area relationship. Bigger areas have more edge, and the edges are where error correction can be escaped more easily, thanks to the very excitability fluctuations that help make the margins marginal. So the more successful patterns tend to be the base from which most variants form.
|

|
So far, so good. But what about those five additional features that
influence the rate of evolutionary change? Happily, we can also
see stability plus the four "catalysts" that speed evolution in this
same neocortical patchwork quilt:
7. Stability may occur, as in getting stuck in a rut (a local minima in the adaptational landscape). Variants occur but they backslide easily. An even background of excitation that avoids the formation of barriers seems a good setup for achieving a stable uniform wallpaperlike pattern. More in a minute. 8. Systematic recombination generates many more variants than do copying errors and the far-rarer point mutations. An end run around a barrier demonstrates one way for variants to met up with clones of their parents. Recombination can occur at those frontiers where different spatiotemporal patterns meet. Though the no-mans-land hexagons, surrounded as they are, are unlikely to start cloning unoccupied territory, the back-and-forth of the frontier leaves behind a composite attractor that could subsequently be activated if the parent active patterns died out in the region. 9. Fluctuating environments (seasons, climate changes, diseases) change the name of the game, shaping up more complex patterns capable of doing well in several environments. For such jack-of-all-trades selection to occur, the climate must change much faster than efficiency adaptations can track it. Fluctuating excitability is highly likely, just from the EEG evidence, and, because its time scale is the milliseconds of the PSPs, it is indeed much faster than the minutes-long LTP mechanisms of neocortex that alter the basins of attraction, sometimes permanently. Thus in cortex, we have the essential setup for evolving elaborate patterns offline, ones that are well in excess of current behavioral requirements. 10. Parcellation, as when rising sea level converts the hilltops of one large island into an archipelago of small islands, typically speeds evolution. As threshold levels rise or background excitation fades, there are many opportunities for chopping up neocortical territories with noncloning barriers. 11. Local extinctions (as when an island population becomes too small to sustain itself) speed evolution because they create empty niches. In the systematic recombination explanation above, there were ghostly patterns in the connectivity, lingering after the active spatiotemporal patterns had died out. Should an active pattern come close to this basin of attraction and start up its spatiotemporal pattern in two adjacent hexagons, it could clone away without any competition for awhile. For a novel pattern, that could represent the chance to "establish itself" -- and some variants.
There are also catalysts acting at several removes, just as in Darwin's example of how cats could improve the clover, and we have no difficulty imagining how this could happen with cortical work spaces. Cloning competitions at the object level could occasionally overrun another competition at the event or metaphor level.
|
|
It is useful to look for biological analogies, for some of the
reasons that I noted at the beginning of the last chapter, but one
shouldn't expect to find perfect parallels. For example, the cortex
may well be the appropriate home for Lamarckism (the pre-darwinian notion that skills acquired during life can be somehow
fed back to the genes, and so inherited by one's offspring). In
cortex, after all, there are generations of lateral copying attempts,
and their improved versions can feed back into the connectivity
changes to influence what happens tomorrow when another
copying attempt is made. In biology, what's wrong with
Lamarckism is the Weismannian genotype-phenotype barrier, the
difficulty of making chimeras, and a distinct individual that
survives as a cohesive unit. So far, the cortex only has a copying
unit (the hexagonal activity doesn't always live and die as a unit,
though embedded attractors encourage that) and it readily
produces chimeras with hybrid vigor in a most unbiological way. You can, of course, ask: Does capture by an attractor constitute stabilizing selection? Does the hexagon correspond to an allele of a gene? A genome (one average organism's worth of genes)? Even a population can potentially be a unit of selection in biology. But here, much as I like finding traditional parallels, I suspect that we are better off focusing instead on the neural substrate -- such as the geometry, chaos, and connectivity concepts of the last few chapters -- rather than seriously seeking parallels in other levels of biology. We have profited from importing island biogeography concepts, even if our hot-spot islands also grow via dendritic amplification mechanisms and raise their sea levels via AGCs. So, too, we may profit from asking about clades, neutral evolution, evolutionarily stable strategies, and (coming up in chapter 8) sexual selection. But we shouldn't expect an exact mapping of either the immune system or cortical darwinian dynamics onto the individuals-that-copy-and-die framework that biological evolution has settled into. Yes, they all feature the darwinian process -- yet the darwinian process isn't really an analogy: it's a crank for complexity that can be turned by instability, whenever a mechanism exists that implements all six essentials. This process just happens to be a major law of the universe, right up there with chemical bonds as a prime generator of interesting combinations, and one apparently able to run on different substrates, each with their own distinctive properties that may, or may not, correspond to those seen elsewhere.
|
What got Hebb to thinking about cell-assemblies was the problem of perception and thought. When, in 1938, Lorente de Nó suggested how cerebral cortex could generate sustained activity with reverberating circuits, he opened up some new possibilities, ones that had seemed closed since Sherrington's day. Hebb wanted a way to think about reactivating a memory trace by thought, without a new sensory stimulus interceding. Lorente's idea gave him sustained activity (the first reason he needed chains of many neurons), but Hebb was still worried about how objects seen from different perspectives were nonetheless perceived as the same object. He considered it a failing of his 1945-1949 cell-assembly theory that it couldn't handle such a key property of concepts.
But what happened was not that the image became sharper. Instead, the subject reported that the image was incomplete; parts of it had a mysterious tendency to fade away and then reappear. In a series of six presentations, controlled by turning on a little light bulb that was also getting a free ride from the eye's movements, the triangle might lose one or another of its three sides. Squares had similar problems, and even faded out entirely on some of the presentations. Images of a face would sometimes lose a nose or eye. I seem to remember, in the excitement over this in the early 1960s (one of the reasons that I switched about then from physics into neurophysiology), that someone claimed to have finally seen a Cheshire Cat doing its disappearing act.
Hubel and Wiesel's orientation-sensitive cortical neurons, found about the same time, suggested an obvious mechanism. Together with the Cheshire Cat fade outs, they resolved the theoretical failing that Hebb identified. It also gave him another reason why a cell-assembly of multiple cortical neurons would be needed: to handle all the line orientations that form the outline of an object. In retrospect, this committee of feature detectors was the most important reason for a cell-assembly. Lorente's reverberating circuits may have gotten Hebb to thinking about neuronal ensembles about 1940, but they really aren't needed to get sustained activity -- something we only learned about in the early 1960s, once we were able to control the firing rate of a single neuron during intracellular recording. All of Hebb's arguments and evidence (summarized in his 1980 book, Essay on Mind) carry over to my more restricted notion of a hexagonal cell-assembly. My triangular arrays add quite a bit of redundancy but their interdigitation also serves to "compress the code" into a half-mm hexagonal space. Such concentrating of the elements allows for lots of little local neural circuits to form, and perhaps change some synaptic strengths in a long-lasting manner. Hebb's dual trace memory is thereby implemented: any adjacent pair of these hexagonal circuits could later reconstitute the firing pattern in all of the multiple triangular arrays that were originally formed during the original stimulus presentation. Hebb unfortunately died in 1985, well before I figured this out. He might have liked how his dual memory traces, his synapses strengthened by success, and his cell-assemblies all came to theoretically hang together in a little hexagon.
|

 Experimental work in Hebb's own lab in Montreal about 1960
eventually provided evidence for various specialized groups
participating in what seemed to be a unitary percept, such as that
of a triangle or square. It was a surprise finding from quite a
different experiment. The physiological tremor of the eyeball is
called micronystagmus; it causes a light-dark boundary in the
image to sweep back and forth over a band as wide as a half-dozen photoreceptors. That can potentially smear the image quite
a bit. You can, however, force the image to move with the eye by
an ingenious system of mounting
the square or triangle just in front
of a contact lens. Whenever the eye
moves, so does its target -- and so
the image is stabilized on the
retina.
Experimental work in Hebb's own lab in Montreal about 1960
eventually provided evidence for various specialized groups
participating in what seemed to be a unitary percept, such as that
of a triangle or square. It was a surprise finding from quite a
different experiment. The physiological tremor of the eyeball is
called micronystagmus; it causes a light-dark boundary in the
image to sweep back and forth over a band as wide as a half-dozen photoreceptors. That can potentially smear the image quite
a bit. You can, however, force the image to move with the eye by
an ingenious system of mounting
the square or triangle just in front
of a contact lens. Whenever the eye
moves, so does its target -- and so
the image is stabilized on the
retina.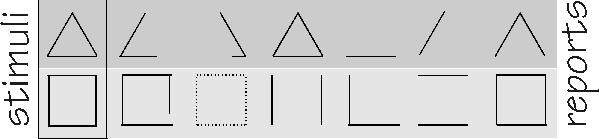
|
Just as the first act introduces the cast and poses the problems,
so the second act tends to show the surprising consequences. My
second act is about the possible products of a fast-acting darwinian principle in neocortex, things such as categories, metaphors,
good guesses, and the train of thought. I'll start with Hebb's
triangle generalization problem and some static-seeming superpositions of schemas, but it's all dynamic underneath: keep
James's train of thought in mind, that series of mental states that
preceded your current one, each one fading into the background
but overlain on its predecessors -- and all capable of contributing
to what connections you're likely to make right now.
But such histories can also be distracting, and we often try to let them fade, try to avoid reexciting them with further thinking. There are various mind-clearing techniques (I take half-hour naps, myself). My friend Don Michael suggests that forming large quasi-stable hexagonal territories might be what meditation with a mantra is all about, preempting the everyday concerns that would otherwise partition the work space and plate out new short-term attractors. By replacing it all with the mantra's nonsense pattern, and holding it long enough for neocortical LTP to fade, the meditator gets a fresh start (for things other than the mantra!). Sounds good to me, but you might forget your grocery list if you carry it too far. An ordinary mantra won't, of course, wipe the work space clean: to prematurely erase those fading attractors, you'll need a fancier mantra that disrupts instead. Short of fogging with seizures, as in electroshock therapy, I don't know of any such eraser schemas -- though one can imagine mental viruses that might preempt entry into those fading basins of attraction, more analogous to an obscuring coat of paint than to a true eraser.
|
|
A neurophysiological theory of higher intellectual function is
the focus of the rest of this book. Ultimately, it's about
consciousness (as the currently dominant patch of the quilt!).
Though you'll never encounter such a list in most of the popular
and academic books about consciousness, any such theory is
going to need to account for: How items of our vocabularies are represented. How memories are stored and recalled. How darwinian shaping-up takes place. How "new ideas" arise, perhaps as pattern variants. Those four, at least, have pretty much fallen out of our search for the darwinian essentials. The rest are harder: The existence of hallucinations and dreams. Déjà vu experiences. Abnormally widespread cloning of an input pattern (perhaps due to lack of competition) might produce the conscious experience usually associated with strong memory resonances that allow widespread cloning. Jamais vu unfamiliarity with the familiar would also be nice to explain. Unreliable memories. Because the long-term synaptic connectivities can be modified by a new active pattern, this could often happen; to avoid this rewriting of history, the subcortical regulation of the cortical competitions probably includes giving permission for the modification of connectivities. Similarly, we'd like something appropriate for concrete thinking and the idée fixe. How abstractions and categories are represented and created. In particular, a theory of mind needs structures suitable for schemas, scripts, syntax, and metaphors. How the various connotations of a word such as "comb" are linked, given that they're likely stored in different cortical areas. The presence of specialized cortical regions that can also participate in nonspecialist tasks. The ability to hold a behavioral set, after selecting it from among possibilities. What might an agenda look like? Subconscious shaping up, while one's attention seems directed elsewhere. Creation of efficient subroutines that function outside of conscious processing, something suitable for Zen archery. A "well-grooved" pattern might "play" from a small cortical territory, like the roll for a player piano, to tie a necktie without competing in the playoffs for consciousness. We do, after all, occasionally manage to do two things at the same time. Serial-order specialties for language and speculative planning, not to mention all the small muscle sequencing that a child needs to tie a shoelace. Speed-of-thought correlates, mechanisms that could vary from time to time in the same individual. During the transitions of manic-depressive illness, a person can go from a fluidity of making connections and decisions to a slow, labored train of thought that lingers too long and fails to make obvious connections. And back again. Our passion for discovering patterns. In our first year of life, we discovered phonemes within words. A year later, we were busy discovering schemas and syntax within sentences, and then we went on to discover narrative principles among more extended discourses. The impression of a narrator, juggling decisions, and speculating about tomorrow. Any explanation needs to be consistent with the neurological evidence that no partial cortical lesion abolishes the "self." That's the kind of coverage needed for a useful theory of mind. It may not have to explain all of two centuries of neurology, one century of psychology, and a half-century of neurobiology and cognitive neuroscience -- but it can't be truly inconsistent with any of it. A theory of mind needs a lot of explanatory power, while still being specific enough to make experimental predictions.
|
|
Theoretical edifices have held some surprises in evolutionary
theory, quite aside from all the emergent phenomena. For
example, one is accustomed to thinking that, if a piece of the
foundation fails, the whole edifice collapses and should be
abandoned. But that isn't necessarily true when dealing with
robust processes such as Darwin's; one can be wrong about
elements of the foundation (as Darwin himself was about
inheritance in 1859, as Clerk Maxwell was in 1865 about the ether)
and still gain many insights about superstructure. Darwin didn't know about Mendel's genes; he thought in terms of blending inheritance, not the particulate inheritance that went on to find its basis in DNA segments called alleles. Although I will use triangular arrays from recurrent excitation and their resulting hexagons as my examples in the coming chapters about the cognitive implications of darwinism in the brain, remember that other mechanisms might prove to be the foundation of what is copied with inherited variation in our neocortex. I'm trying to piece together a general theory for how the superficial layers of neocortex could run the darwinian ratchet, but I'm also -- in Act II -- carrying on William James's project, attempting to show how any such darwinian theory could account for higher intellectual function. As I give examples of how categories could be created, and elaborated into schemas and metaphors, try to distinguish the emergent structures per se -- and perhaps guess how they could be implemented with different darwinian building materials, ones that still satisfy the six essentials and provide a few accelerating factors in the manner of my triangular arrays and hexagonal mosaics.
|

Act III suggest that memory is organized with a framework of motor programs within the brain. Far from being a black box of no relevance to behaviorist psychology, the brain is a jack-in-the-box, filled to the brim with spring-loaded plans of action. As such it is the very wellspring of all behavior. These brain-mind behavior programs, like sensory representations, are virtual — and they have even been called "fictive" to capture their promissory aspect — but they are no less real. In our being and becoming, they are us and we are they. |
 You are reading THE CEREBRAL CODE.
You are reading THE CEREBRAL CODE.
The paperback US edition |