|
William H. Calvin |
|
Calvin, W.
H. (1987). The brain as a Darwin
machine. Nature 330:33-34 (5 November). Available at
WilliamCalvin.com/1980s/1987Nature.htm AMIDST all the hyperbole about thinking machines that has accompanied the emergence of large-scale parallel computers from their serial predecessors, we have begun to contemplate the prospect of simulating some of our brain's massive parallelism. But one immediately runs into a role reversal worthy of a Mozart opera: the most distinctively human higher brain functions are surprisingly serial. Human beings are perpetually stringing things together: phonemes into words, words into sentences, concepts into scenarios - and then fussing about getting them in the right order. Our brain uses word-order rules to create a very productive language, with an infinite number of novel messages, rather than the several dozen standard interpretations Dissociated with the several dozen cries and grunts of any other primate species. It is not our mellifluous voices that constitute a significant advance but rather our arrangement rules, the meaningful order in which we chain our utterances..... [The rest is in PDF format.]
|
For parallel computers to simulate our brains, we must face the fact that human beings have a better claim on the title Homo seriatim than Homo sapiens - - we're more consistently serial than wise. |
|
William H. Calvin, "The missing LINC." Byte (April 1982). Available as
PDF. I keep seeing the "personal computer era" referred to as having begun in the mid-1970s, as if it required the microchip to make it possible to design hardware and software for a single-user computer. But there are important antecedents to this, dating back twenty years, and they were not hand-held calculators so much as proper computers with analog interfaces and mass storage-namely, lab computers. In 1962, at MIT's Lincoln Labs, Wesley Clark and Charles Molnar designed the LINC (Laboratory INstrument Computer), to be used in a research lab in a manner analogous to an oscilloscope. It wasn't merely its display and analog-to-digital converters (hence joysticks and Spacewar) which made it unique: its software was designed to enable the scientist-user to program without a professional programming staff. Much of the design rationale was process-control oriented (hence interrupts) so that online data analysis could be performed during an experiment, allowing modification to the experimental protocol. But having such a friendly computer in the room, shared only with the other users of the same lab, created the atmosphere of "personal computing" a decade before mass-market economics extended it. |
 |
|
Calvin, W. H., Devor, M., and Howe, J.
F. (1982). Can neuralgias arise from minor demyelination? Spontaneous firing,
mechanosensitivity, and afterdischarge from conducting axons. Experimental
Neurology 75:755-763.
Available at
WilliamCalvin.com/1980s/1982ExpNeurol.pdf Mammalian peripheral axons respond to local disruption of their myelin sheath with membrane changes which support continuous conduction of the impulse through the affected region. We report here that sites of demyelination may become foci of spontaneous impulse initiation. Such sites may also generate ectopic discharges upon slow mechanical distortion. Finally, conduction of an impulse train through a demyelinated region may set off an ectopic afterdischarge that may last many seconds. Rhythmic ectopic firing in dysmyelinated but conducting axons is very similar to that observed in regenerating axons and nerve-end neuromas. Although the latter have long been recognized as sources of pathophysiologic sensations, this is the first indication that neuralgias could arise following minor dysmyelination in peripheral nerves without substantial conduction deficits. |
|
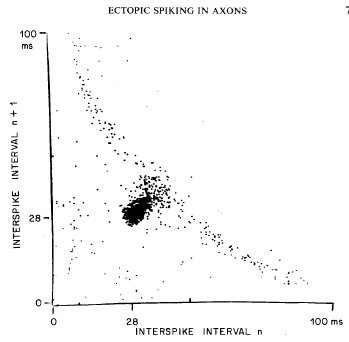
|
Calvin, W. H. (1980). Normal repetitive
firing and its pathophysiology. In: Epilepsy: A Window to Brain
Mechanisms (J. Lockard and A. A. Ward, Jr., eds.), Raven Press,
New York,
pp. 97-121.
Available at
WilliamCalvin.com/1980s/1980epilepsy.pdf A neuron communicates over long distances (more than a few millimeters) by generating a train of impulses which propagates down the axon to release a series of prepackaged quanta of neurotransmitter molecules. The rate, or perhaps the patterning, of the impulse train carries the information. One of the hallmarks of an interictal epileptogenic focus is that many of its neurons are observed to cluster their impulses into bursts, with the intervals between impulses being unusually short (several milliseconds). Is the bursting neuron some sort of pacemaker, driving other normal neurons into synchronous activity and thus spreading the trouble? Or is the bursting one observes just one of those recruited neurons, having nothing more wrong with it than an oversized synaptic input? Or perhaps there are no pacemaker neurons; the trouble could be subtly distributed over many neurons, changing the balance of excitation and inhibition so that the whole circuit tends to go into a bursting-type oscillation. |
|
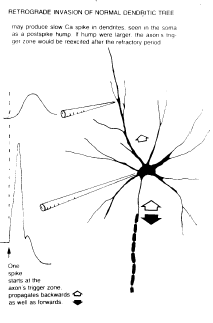
|
Calvin, W. H., and Graubard, K. (1979). Styles of neuronal computation. Chapter 29 in: The Neurosciences, Fourth Study Program. Edited by F. O. Schmitt and F. G. Worden. Cambridge: MIT Press, pp.513-524. Available at WilliamCalvin.com/1970s/1979StylesNeurocomputation.pdf Can be viewed and printed in PDF format ABSTRACT Neurons seldom serve as relay stations; they typically transform their inputs in some manner to produce a new output function. A conventional processing path within a neuron includes a cascaded series of steps, each capable of contributing characteristic styles of computation: passive spread of PSPs to a spike trigger zone, spike initiation and repetitive firing, spike propagation, and ultimately synaptic output. We now know of many neurons where this one-way cascade must be supplemented with additional processes, such as intermittent conduction or dendritic spikes. There are also simpler cases, such as the passive-to-synaptic cascade of spikeless neurons. Styles of computation (e.g., arising from transient or sustained responsiveness, from high or low thresholds, and so forth) may be contributed by different stages of the cascade (e.g., from spike initiation regions, from conduction in axons, from synaptic input-output processes). Processing may thus differ among the various presynaptic regions of the same neuron. |
|
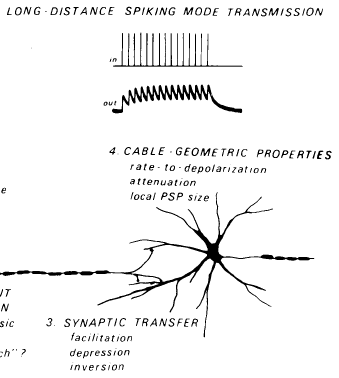
|
Graubard, K., and Calvin, W. H. (1979).
Presynaptic dendrites: Implications of spikeless synaptic transmission and
dendritic geometry. Chapter 18 in: The Neurosciences, Fourth Study Program.
Edited by F. O. Schmitt and F. G. Worden.
Cambridge: MIT
Press, pp.317-331.
Available at
WilliamCalvin.com/1970s/1979PresynapticDendrites.pdf voltage. In a number of invertebrate spikeless and spiking neurons, synaptic transmission lasts for the duration of the presynaptic depolarization. There is a threshold presynaptic voltage, but it is sometimes below the spike threshold, or even below the resting potential. Above the release threshold, the postsynaptic potential (PSP) grades over a wide range of presynaptic voltages. The steady-state cable equation can be used to predict how PSPs spread within a complex dendritic geometry. Input synapses located on long, thin processes develop large PSPs, although these PSPs attenuate markedly before reaching central structures. Whereas a proximal dendrodendritic output synapse would see proximal and distal input synapses as approximately equal, a distal output synapse would see many-fold differences in the relative PSP sizes. Thus regional computations could take place in distal dendritic trees ; a given neuron could compute many different functions of' its inputs. Models are made of Aplysia and lobster neurons, of a cat spinal motoneuron, and of a neuron in the rat superior colliculus. 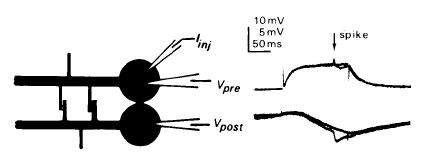 |
|
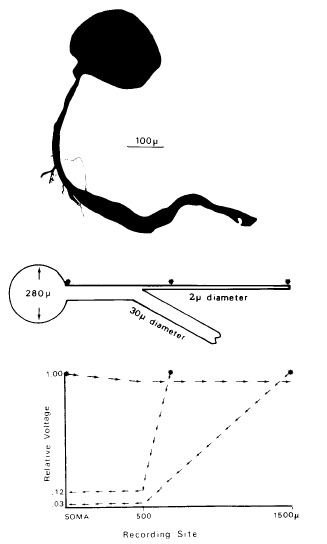
|
Calvin, W. H., and Hartline, D. K. (1977). Retrograde invasion of lobster stretch receptor somata in the control of firing rate and extra spike patterning. Journal of Neurophysiology 40:106-118. Available at WilliamCalvin.com/1970s/1977JNeurophysiol.pdf Can be viewed and printed in PDF format SUMMARY AND CONCLUSIONS |
|
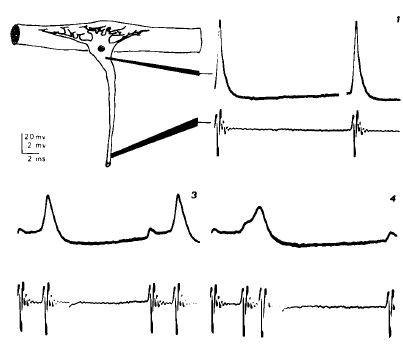

|
Calvin, W. H., and Sypert, G. W. (1976). Fast and slow pyramidal tract neurons: An intracellular analysis of their contrasting repetitive firing properties in the cat. Journal of Neurophysiology 39:420-434. Available at WilliamCalvin.com/1970s/1976JNeurophysiol.pdf Can be viewed and printed in PDF format SUMMARY AND CONCLUSIONS1. Intracellular recordings were made from an estimated 500 neurons in the sensorimotor cortex of barbiturate-anesthetized cats. Of those which were antidromically identified from the medullary pyramids, 70 were selected which also exhibited steady repetitive firing to steps of current injected through the recording electrode; 81% were "fast" (conduction velocity greater than 20 m/s) and 19% were "slow." 2. As shown by earlier workers, the spike duration is a function of conduction velocity ; a spike duration of 1.0 ms is the dividing line between fast and slow. 3. Of the 57 fast pyramidal tract neurons (PTNs), 14 exhibited double spikes during otherwise rhythmic firing patterns to a step of injected current. These very short interspike intervals (usually 1.5-2.5 ms) were first seen interspersed in a rhythmic discharge (e.g., 50-ms intervals) but, with further increases in current strength, would come to dominate the firing pattern ; e.g., double spikes every 40 ms. Further increases in current would typically shorten only the long intervals ; e.g., 40-30 ms, but some fast PTNs developed triple spikes, etc. 4. The extra spike appears to arise from a large hump which follows most spikes in fast PTNs; while this humplike "depolarizing afterpotential" can also be seen in slow PTNs, it is small. Extra spikes were seen only in fast PTNs with large postspike humps ; in perhaps half of the fast PTNs, extra spikes probably contributed to "adaptation." 5. Slow PTNs often had frequency-current curves which were not repeatable ; a "hysteresis" phenomenon could often be seen, where the proportionality constant relating current to firing rate decreased following high firing rates. 6. The B spike was distinguishable from the A spike in differentiated antidromic spikes in 77% of the slow PTNs, in only 14% of the fast PTNs which later exhibited double spikes during current-induced repetitive firing, and in 53% of the other fast PTNs. 7. The antidromic spike heights of doublet PTNs were not significantly different from those of other repetitively firing PTNs. |
|
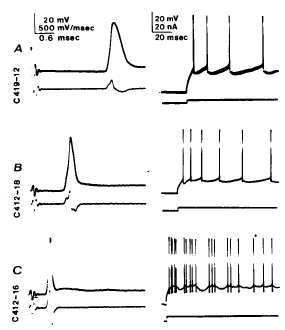
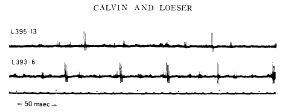
|
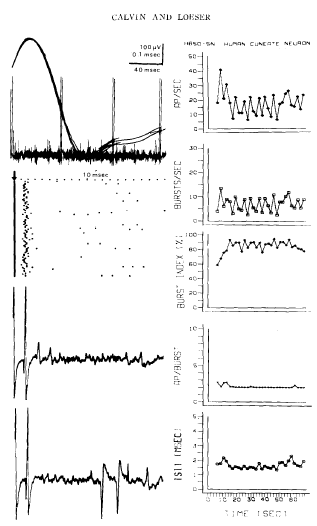
Calvin, W. H., and Loeser, J. D. (1975). Doublet and burst firing patterns within the dorsal column nuclei of cat and man. Experimental Neurology 48:406-426. Available at WilliamCalvin.com/1970s/1975ExpNeurol.pdf - Can be viewed and printed in PDF format in two parts: Part 1 Part 2 We have examined extracellularly the firing pattern of neurons in the cat external cuneate nucleus and in the human main cuneate nucleus, focusing upon both the spontaneous firing patterns and its modification by natural stimulation. Many of these neurons exhibit stereotyped doublet or burst firing patterns, e.g., the interval between the spikes might be 1 .0 + 0 .1 msec in a given cell . For most cells, this characteristic doublet interval was between 0.8 and 2.0 msec, with a few extending to 5 msec . While doublets were most common, the number of spikes per burst ranged to six or more . When the external cuneate neurons were synaptically driven by forelimb position changes, the firing rate increased but the proportion of spikes occurring within bursts (the "burst index") often fell . The doublets that occurred became broader during synaptic drive. This paradoxical behavior (the peak instantaneous firing rate falling as the average firing rate rises) is analogous to the doublet firing patterns occasionally observed in spinal motoneurons ; there, the doublet is caused by a large depolarizing afterpotential (postspike hump) which rises through the falling threshold at the end of the relative refractory period to elicit an "extra spike ."
|
|
|
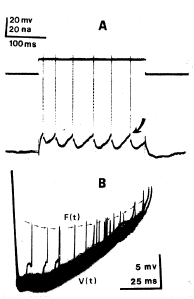
Calvin, W. H. (1974). Three modes of repetitive firing and the role of threshold time course between spikes. Brain Research 69:341-346. Available at WilliamCalvin.com/1970s/1974BrainRes.pdf - Can be viewed and printed in PDF format When rhythmic firing is elicited in cat spinal motoneurons by long-lasting depolarizing currents, the `threshold' voltage from which each spike arises may appear to be quite constant (Fig. IA, arrow). If one probes for the threshold during the interspike interval (ISI), however, it is seen to fall well below this level, rising towards it later in the ISL Exceptionally large depolarizing afterpotentials may intersect this threshold time course shortly after a spike, causing an extra spike. The extra spike itself may also similarly produce another extra spike; thus a regenerative cycle may produce a burst of spikes at a high firing rate. |
|
|
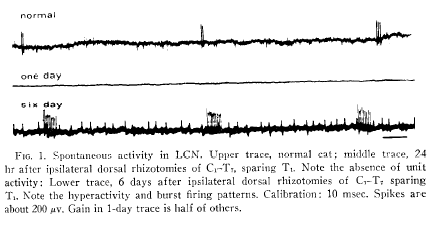
TERRELL D. KJERULF, JOHN T. O'NEAL, WILLIAM H. CALVIN, JOHN D. LOESER AND LESNICK E . WESTRUM
Deafferentation Effects in Lateral Cuneate Nucleus of A correlated anatomical and physiological investigation of the effects of
unilateral cervicothoracic dorsal rhizotomies upon lateral cuneate nucleus of
the cat (LCN) is reported. Pairs of adult cats with identical survival times
were selected to correlate structural and functional changes . Two phases are
described in the development of alterations of neuronal firing patterns. In the
first phase, a relative silence within LCN was associated with depletion of
round synaptic vesicles in the presynaptic profiles (LR boutons) of primary
dorsal root afferents . The second phase was characterized by a development of
spontaneous electrical hyperactivity which corresponded anatomically to the
presence of denuded postsynaptic specializations, transient increase of adjacent
extracellular space and an apparent decrease in the number of dendritic spines.
There was a persistence of an unaltered population of small presynaptic boutons
with flattened vesicles (SF boutons) . The LCN neuronal membrane is viewed as
having an intrinsic tendency for repetitive firing which is enhanced by the
functional |
 |
|
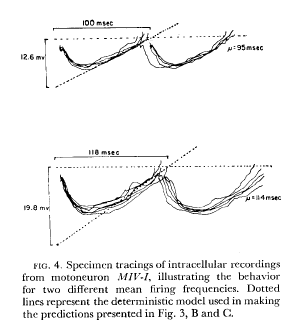
Calvin, W. H., and Stevens, C. F. (1968). Synaptic noise and other sources of randomness in motoneuron interspike intervals. Journal of Neurophysiology 31:574-587. Available at WilliamCalvin.com/1960s/1968JNeurophysiol.pdf - Can be viewed and printed in PDF format SUMMARY In one class of cells the spike-generator mechanism appeared to be adequately
described by a model consisting of a linearly increasing depolarization that
generates a spike on reaching a constant firing level and is then reset. In a
second class of cell the firing level increased approximately linearly with time
since the last spike, and this feature had to be included in the model of spike
generation In two instances synaptic noise alone was not adequate to account for the observed interspike-interval variability, but further analysis revealed an additional source that would account for the variability. In one case a multimodal histogram appeared to arise from "local responses" that tended to occur at preferred times. In the other instance the rate of depolarization appeared to change from interval to interval ; by including this additional noise source it was possible to account for the observed variability. |
|

|
Calvin, W. H., and Stevens, C. F. (1967). Synaptic noise as a source of variability in the interval between action potentials. Science 155:842-844. Available at http://WilliamCalvin.com/1960s/1967Science.pdf Abstract. The source of variability in the interval between action potentials has been identified in a class of cat spinal motoneurons. The observed random fluctuations in membrane potential (synaptic noise) together with an empirical description of spike generation accurately predict the statistical structure of variability observed to occur in the neuron's discharge.
|
|
|
Ehrlich, A., and Calvin, W. H. (1967). Visual discrimination behavior in Galago and owl monkey. Psychonomical Science 9:509-510. Available at http://WilliamCalvin.com/1960s/1967Galago.pdf Two nocturnal primates, galago and owl monkey, were inferior to rhesus monkeys in ability to detect small differences in brightness. Neither was able to learn a red-green discrimination. Although galagos did somewhat better than owl monkeys on pattern discriminations, both species were inferior to rhesus monkeys. |
|
|
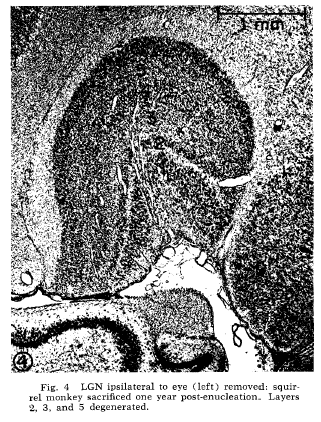
Doty, R. W., Glickstein, M., Calvin, W. H. (1966). Lamination in the lateral geniculate nucleus in the squirrel monkey, Saimiri sciureus. Journal of Comparative Neurology 127:335-340. Available at http://WilliamCalvin.com/1960s/1966JCN.pdf ABSTRACT The parvocellular portion of the lateral geniculate nucleus (LGN) in the squirrel monkey (Saimiri sciureus) is a relatively unlaminated mass of cells. In order to determine whether a concealed laminar structure might exist, one eye was removed in each of three squirrel monkeys. Transneuronal degeneration did not appear prominent in the LGN of the animal which was allowed to survive for six months, but in the two animals which survived for one year six laminae were clearly present. There was transneuronal degeneration in layers 1, 4 and 6 contralateral to the enucleation, and 2, 3 and 5 ipsilaterally. Lack of interlaminar fiber masses probably accounts for the apparent absence of lamination in the normal parvocellular mass.
|
|
|
William H. Calvin and Charles F. Stevens
"A Markov Process Model for Neuron Behavior in the Interspike Interval." Proceeding of the 18th Annual Conference, on Engineering in Medicine & Biology, 1965, 1:118 Available at http://WilliamCalvin.com/1960s/1965MarkoffProcess.pdf
|
|

| xxxx |
xxxx |
| xxxx |
xxxx |
| xxxx |
xxxx |
| xxxx |
xxxx |